
Пневмококки-комменсалы превращают течение опасных инфекционных заболеваний в мирную колонизацию микробиома, управляя ядерной деметилазой KDM6B (фермент, участвующий в воспалительном ответе и альтернативном сплайсинге) клеток хозяина, помечают участки нуклеотидных последовательностей для транскрипционного фактора NF-κB (транскрипционный фактор, контролирующий экспрессию генов, участвующих в иммунном ответе, клеточном цикле, апоптозе) промотора цитокина ИЛ-11, повышая регенерацию эпителия.
С помощью факторов вирулентности, действующих во внеклеточном пространстве и в цитоплазме, бактериальные патогены управляют реактивностью макроорганизма и способствуют заражению инфекционными возбудителями. На что сегодня мало обращают внимание, так это на модуляцию внутриядерных процессов бактериями для контроля иммунологического ответа макроорганизма. В этой статье «Nature Microbiology» Connor с соавт. описывают, что бактерии изменяют эпигенетический ландшафт хозяина, превращая смертельную инфекцию в бессимптомный комменсализм [1].
Бактерии секретируют «нуклеомодулины», которые влияют на транскрипционную активность нуклеосом, воздействуя на эпигенетические модификаторы ядерных белков и ДНК [2]. Некоторые бактерии либо активируют, либо угнетают транскрипцию, изменяя доступность промоторов генов макроорганизма путем ацетилирования или метилирования гистонов, соответственно [3].
Если гены, вовлеченные во врожденный иммунный ответ организма, нацелены на эпигенетическую модификацию, можно предполагать, что течение инфекции будет изменено в пользу патогена, способствуя гипер- или гиповоспалению. Например, сигнализация посредством NF-κB является ключевым путем в процессе воспаления, а для формирования полноценного ответа NF-κB на инфекцию необходима хроматиновая перестройка. Таким образом, «атака на клеточное ядро», изменяющая ремоделирование хроматина, может оказаться очень эффективным способом превратить инфекцию в комменсализм. До сегодняшнего дня было сложно доказать эту гипотезу.
В ходе ранних работ над Helicobacter, Escherichia coli, Campylobacter и Mycobacterium leprae были получены первоначальные доказательства того, что во время бактериальных инфекций возникают эпигенетические изменения в клетках макроорганизма, такие как деацетилирование гистонов [4]. Позже в ходе полногеномного секвенирования обнаружилось, что многие бактерии кодируют белки с SET-доменами [белковые домены, обычно с метилтрансферазной активностью, аббревиатура представляет собой акроним от названий мух-дрозофил, бывших объектами исследований, — прим. перев.], характерными для эукариотических метилаз, которые могут вызывать посттранскрипционные модификации гистонов [5]. Сейчас известно, что бактерии могут эпигенетически модифицировать ДНК и гистоны макроорганизма с помощью нескольких посттрансляционных модификаций, таких как убиквитинирование, сумоилирование (SUMOylation, от англ. s(mall) u(biquitin-related) mo(difier) — разновидность посттрансляционной модификации белка), фосфорилирование, ацетилирование и метилирование [6]. Однако обнаружение эпигенетически-модифицирующих ферментов у бактерий не доказывает, что эти модификаторы функционируют во время болезни. Важным ранним признаком существования такой связи стало наблюдение потери Bacillus anthracis белка BaSET, вследствие чего смертельно опасная сибирская язва превращалась в нелетальную инфекцию [5]. В представленных Connor с соавт. данных показано, что модификация эпигенетического статуса макроорганизма в пользу бактерий лежит в основе запрограммированной модели течения болезни. Удивительно то, что в ходе исследования удалось показать, что эпигенетическая модификация изменяет баланс в организме не в сторону развития патологии, а в сторону колонизации.
Streptococcus pneumoniae, патогенная бактерия для человека, либо колонизирует носоглотку (что выражается в бессимптомном носительстве), либо поражает легкие, кровеносную систему и мозг, становясь причиной значительной смертности. Плохо изучено, что определяет развитие различных штамм-специфичных патологий. Действительно, в большей части опубликованных исследований внимание сосредоточено на развитии патогенеза вследствие инфицирования бактериями, а не на аспекте колонизации. Connor с соавт. изучили транскрипционный ответ макроорганизма в эпителиальных клетках дыхательных путей во время неинвазивного контакта с пневмококками и обнаружили повышенную экспрессию NF-κB, зависящую от эпигенетического модификатора KDM6B макроорганизма. KDM6B представляет собой стиратель посттрансляционных модификаций у эукариот, который деметилирует лизин (в 27-й позиции) гистона H3. Используя метод секвенирования иммунопреципитации хроматина, авторы показали, что KDM6B специфически ремоделирует промотор интерлейкина (ИЛ)-11 для обеспечения уникального паттерна связывания p65, что приводит к увеличению продукции этого цитокина (см. рис. 1).
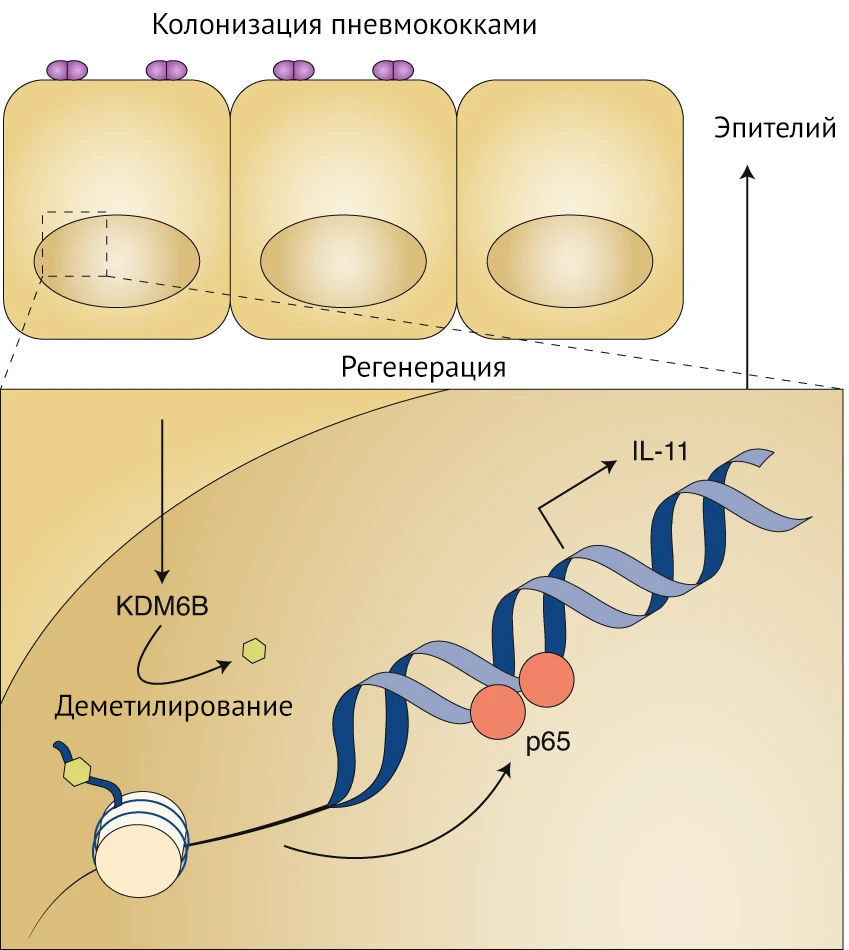
Рисунок 1 | Пневмококки-комменсалы изменяют эпигенетику макроорганизма для увеличения выработки ИЛ-11 и сохранения целостности эпителияВ противоположность инвазивным штаммам, пневмококки, бессимптомно колонизирующие носоглотку, активно вызывают эпителиальную регенерацию. Деметилаза KDM6B гистона Н3 удаляет метильную модификацию с лизина-27 в промоторе ИЛ-11. Это, в свою очередь, увеличивает связывание p65 для индукции ИЛ-11 и поддерживает целостность и восстановление эпителия.Затем они исследовали, как ИЛ-11, один из цитокинов, необходимых для заживления ран, изменяет физиологию макроорганизма в сторону развития бессимптомной колонизации, а не летального инфекционного заболевания. Было выявлено, что колонизирующие штаммы, такие как серотипы 6B, 19A и 19F (Str. pneumoniae), индуцировали уникальную перестройку промотора ИЛ-11 с помощью KDM6B, вследствие чего увеличивалась продукция ИЛ-11. Это, в свою очередь, приводило к меньшему повреждению эпителия по сравнению с инвазивными штаммами, такими как серотипы T4 и 1. Удалось также обнаружить, что химическое ингибирование KDM6B способствует переключению программы колонизации штамма пневмококка 6B на инвазивное заболевание, а введение в экспериментальную модель экзогенного ИЛ-11 превращало инвазию штамма T4 в колонизацию.
В ходе исследования Connor с соавт. также выявили, что представители нормального микробиома носоглотки, такие как стрептококки группы D, Eikenella и Fusobacterium, используют один и тот же человеческий генетический стиратель, чтобы сформировать эпигенетический ландшафт мирного сосуществования, связанного с присутствием ИЛ-11. Обобщая, можно полагать, что бактерии, в течение длительного времени колонизирующие слизистую оболочку, активно и, возможно, оказывая стимулирующее воздействие, вызывают клеточную регенерацию и репарацию у макроорганизма. Эти процессы снижают степень повреждения слизистой оболочки путем уникальной посттрансляционной модификации для модулирования неканонической подгруппы мишеней активации NF-κB.
Как и во всех неожиданных поворотах науки, вопросы, возникающие в результате этих экспериментов, крайне любопытны. Какой бактериальный фактор взаимодействует с KDM6B? Полезны ли эти факторы также для метаболизма бактерий, как следует из их влияния на скорость роста [1]? Почему деметилирование гистона H3 с помощью KDM6B влияет только на часть общих генов-мишеней NF-κB? Требуется ли для поддержания мирной колонизации тонизирующая активность при восстановлении с помощью ИЛ-11? На перспективу использования новых методик терапевтического вмешательства указывает успех применения семейства ингибиторов гистондеацетилаз в лечении рака и других хронических заболеваний [7]. Может ли быть так, что активация другой модификации (например, деметилирование гистонов) представит новую возможность ослабить воздействие инфекции там, где ингибиторы гистондеацетилазы оказались бессильны [8]? Прямое деметилирование ДНК является основой рестрикционной модификации в битве между бактериями, и оно может оказаться еще одним уровнем атаки бактерий на ядро клеток макроорганизма.
Ясно, что бактерии научились проникать сквозь ядерную мембрану клеток человека; остается надеяться, что исход этого взаимодействия будет благоприятным для нас.


