

Гепатит А — острое инфекционное заболевание печени, вызываемое вирусом гепатита А (англ. HAV). Заражение HAV может приводить к развитию смертельно опасного молниеносного гепатита (острой печеночной недостаточности). С «болезнью немытых рук» близко знакомы миллионы людей, а вот HAV пока остается одним из самых малоизученных вирусов. Недавно была получена кристаллическая структура этого инфекционного агента. Удивительно, но она оказалась похожей на структуру пикорнаподобных вирусов насекомых. Возможно, эта информация сможет пролить свет как на происхождение HAV, так и на некоторые его загадочные свойства.
По данным ВОЗ, в мире ежегодно регистрируется около 1,4 млн случаев заболевания гепатитом А. В основном это удел населения развивающихся стран с неблагополучной санитарной обстановкой и низким уровнем личной гигиены [1]. Но именно в таких странах проживает большинство землян. Почему же до сих пор возбудитель столь распространенной и опасной инфекции не был досконально изучен? Одна из причин — его необычная структура. Вирус гепатита А относится к семейству Picornaviridae. Пикорнавирусы — маленькие икосаэдрические вирусы, у которых нет внешней оболочки, а геном представлен одноцепочечной РНК. К пикорнавирусам относятся одни из самых маленьких известных патогенов человека и животных (рис. 1), в том числе такие привычные нам «простудные» вирусы [2].
Структура пикорнавирусов
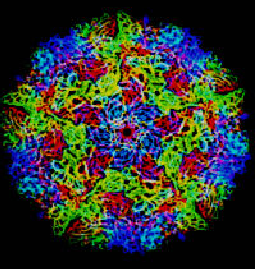
Все вирусы имеют белковую оболочку, предохраняющую их нуклеиновую кислоту от внешних воздействий — капсид (от лат. capsa — вместилище, ящик). Капсид, в свою очередь, состоит из отдельных одинаковых структурных единиц (белков) — капсомеров. Капсид пикорнавирусов имеет шарообразную структуру диаметром 30 нм и состоит из 60 копий каждого из четырех структурных белков — VP1, VP2, VP3 и VP4. Кристаллическая структура пикорнавирусной частицы следующая: белки VP1, VP2 и VP3 состоят из 240–290 аминокислотных остатков; каждый из белков принимает форму восьмицепочечной антипараллельной β-складчатой структуры (так называемая структура «рулет с вареньем» (jelly roll)) (рис. 2). Эти белки образуют наружную поверхность капсида, толщина которого составляет приблизительно 5 нм. Белок VP4 состоит всего лишь из 70 аминокислотных остатков и лежит поперек внутренней поверхности капсида [3]. В итоге капсид пикорнавирусов приобретает форму икосаэдра, или, другими словами, он обладает икосаэдрической симметрией. То есть субъединицы вируса расположены в виде правильного икосаэдра вокруг РНК, скрученной в клубок. Белковые субъединицы (капсомеры) больших икосаэдрических вирусов для сохранения формы сгруппированы в морфологические группы, различимые с помощью электронного микроскопа: это могут быть пентамеры (5 субъединиц) или гексамеры (6 субъединиц), формирующие в капсиде каналы [4, 5]. Структурный белок VP1 образует пентамеры, к которым примыкают белки VP2 и VP3, образующие вершины икосаэдра с трехлучевой симметрией. Между пентамерами и белками VP2 и VP3 образуется углубление, которое называется каньон. В этом каньоне у большинства пикорнавирусов есть так называемый карман — пространство, в которое при упаковке вируса попадает небольшая молекула, обычно некий липид. Этот липид, или «карманный фактор», нужен для того, чтобы стабилизировать капсид вне клетки. Как только вирус связывается с нужным рецептором и попадает в клетку, фактор выходит из кармана, запуская распаковку частицы и делая возможным проникновение РНК в цитоплазму [2].
Сверчок и HAV: связь раскрыта!
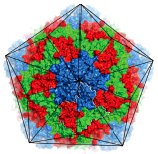
Как выяснила группа ученых из Великобритании и Китая, структура вируса гепатита А отличается от таковой других пикорнавирусов [6]. Внешняя поверхность HAV — гладкая, без углублений (рис. 3). Основные белки капсида — VP1, VP2 и VP3 — формируют восемь антипараллельных β-структур и охватывают всю толщину капсида. В целом, HAV является высокоупорядоченным вирусом, и организация его белковых цепей сходна с таковой других представителей Picornaviridae. Однако ученые нашли несколько отличий. Во-первых, у вируса гепатита А отсутствует типичный для пикорнавирусов «карманный фактор». Вторая тонкая деталь — небольшое изменение в N-конце белка VP2, а именно — небольшое сгибание на границе пентамеров. Это сгибание усиливает взаимодействие одного пентамера с соседним за счет образования дополнительной цепи на VP2-β-структуре. В итоге меняется связность субъединицы: смежные протомеры (полипептидные цепи, являющиеся исходной единицей организации структуры белка) одного пентамера сшиты вместе примыкающим пентамером. Из всех пикорнавирусов такой структурой обладает только вирус гепатита А, однако похожие структуры есть и у пикорнаподобных вирусов насекомых, например, у паралитического вируса сверчка (cricket paralysis virus (CrPV)). Таким образом, некоторые загадочные свойства HAV могут объясняться тем, что этот вирус является связующим звеном между «современными» пикорнавирусами и более «примитивными» предшественниками вирусов насекомых.
Новые факты о HAV
Было обнаружено, что вирусные частицы HAV более устойчивы к физическим и химическим воздействиям, чем частицы других Picornaviridae [6]. Они остаются стабильными при температуре до 80 °С и значениях pH около 2. Можно было бы предположить, что такая устойчивость вызвана изменениями домена VP2 на границе пентамеров, что описывалось выше. Однако, стабильность паралитического вируса сверчка, организованного подобно HAV, ничем не отличается от таковой прочих пикорнавирусов. Дальнейшие исследования показали, что устойчивости частицы HAV способствует состыковка небольших аминокислотных остатков и тирозиновых боковых цепей, прижатых вокруг оси двойной симметрии.
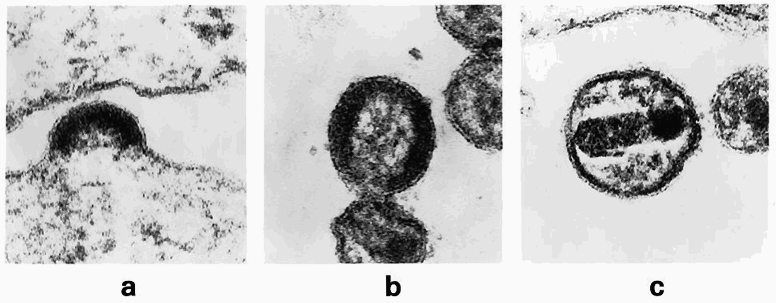
Также HAV отличается от других пикорнавирусов (в частности, энтеровирусов) и механизмом проникновения в клетку-хозяина. Энтеровирусы передают свой геном через поры в мембране эндосомы, формируемые амфипатической (включающей участки с противоположными — гидрофильными и гидрофобными — свойствами) N-концевой спиралью VP1 и белком VP4. Процесс инфицирования состоит из двух шагов: проникновения вирусной частицы и высвобождения ее генома. Первый шаг для HAV хорошо изучен: вирус без изменений попадает в клетку путем трансцитоза (что как раз-таки свойственно пикорнаподобным вирусам насекомых), тогда как второй шаг — высвобождение РНК-генома — остается неясным. Возможно, для разборки вирусной частицы требуются специфические факторы инфицируемой клетки-хозяина. Недавние исследования показали, что HAV содержит тандем мотивов YPX3L, которые связывают ALIX-компонент ESCRT-пути [7]. То есть вирус способен привлекать ESCRT-комплекс, обычно служащий для облегчения высвобождения частиц оболочечных вирусов путем экзоцитоза. ESCRT (endosomal sorting complex required for transport) — совокупность белков, необходимых для мембранных перестроек при формировании выпячиваний и впячиваний. Работая вместе с другими вспомогательными белками, ESCRT включает режим ремоделирования (изменения) мембраны, что приводит к ее изгибанию, отделению от цитоплазмы и дальнейшему «перерезанию» шейки выпячивания. Таким образом происходит выход вируса, или вирусный бадинг (англ. budding — почкование) (рис. 4). ESCRT играет важную роль в ряде клеточных процессов, таких как биогенез мультивезикулярных тел, отделение клеток и попадание вирусов в клетку. Чтобы помочь вирусу гепатита А проникнуть в клетку, белок ALIX связывает и рекрутирует ESCRT-III, компоненты которого участвуют в основных процессах ремоделирования мембраны.
HAV — переходная эволюционная форма
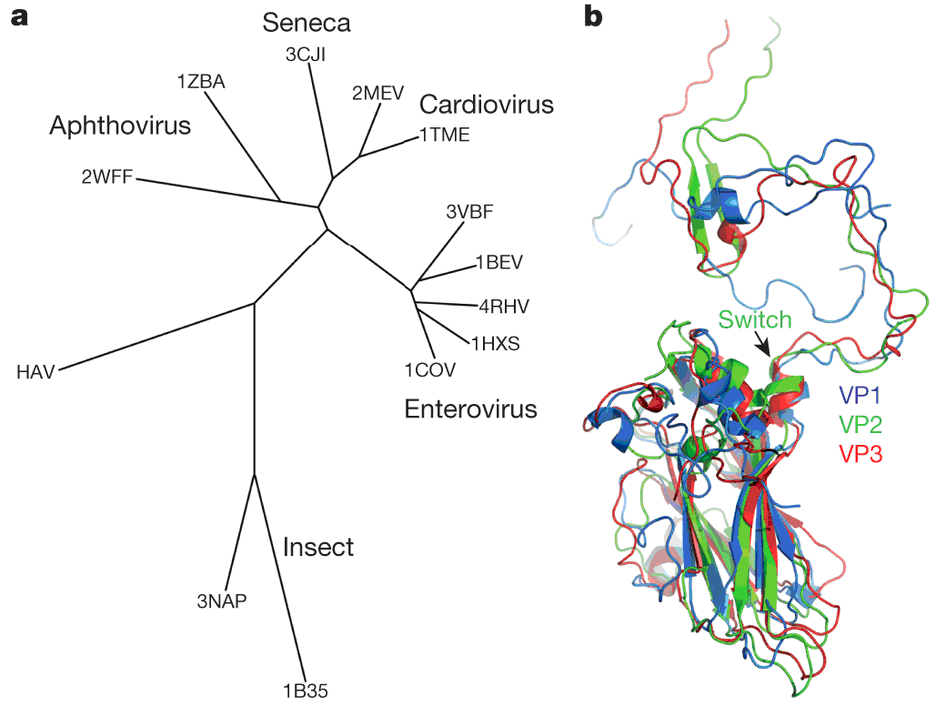
Таким образом, можно сделать вывод, что HAV обладает тонкими, но глубокими структурными различиями с другими пикорнавирусами. Филогенетический анализ показывает, что HAV связан как с классическими пикорнавирусами, так и с пикорнаподобными вирусами насекомых (рис. 5а). Кроме того, замена N-концевого домена в белке VP2 повышает уровень его гомологии с белками VP1 и VP3 (рис. 5b). Это подтверждает идею о том, что HAV сохранил структурные и функциональные особенности первичных пикорнавирусов, которые были родственны пикорнаподобным вирусам насекомых. Та же замена в белке VP2 позволила вирусу гепатита А приобрести эффективные механизмы внедрения своего генома в клетку, что стимулировало взрывное появление разнообразных форм современных пикорнавирусов млекопитающих.
Полученные знания о кристаллической структуре и происхождении HAV раскрывают широкие перспективы в борьбе с этим вирусом. Например, можно синтезировать препараты, направленные на разрушение структуры вируса. Или, зная, что HAV, как и его предки, попадает в клетки путем трансцитоза, пытаться блокировать этот процесс. Можно сделать лекарства, направленные на разрушение комплекса HAV-ESCRT, и таким образом предотвращать активную репликацию вируса в клетках. Думаю, что исследования в этой области не заставят себя ждать, и скоро мы узнаем о появлении на рынке новых препаратов, способных эффективно бороться с вирусом гепатита А.
Источник: Биомалекула


